
Gentechnik Impfungen – Wohin geht die Reise?
Der folgende Beitrag wurde von Dr. Sucharit Bhakdi, Dr. Karina Reiss und Dr. Michael Palmer am 5. November 2022 auf www.globalresearch.ca veröffentlicht.
Hier eine Übersetzung ins Deutsche.
Beweggründe für die Entwicklung von Impfstoffen
Das Konzept, das der Entwicklung von Impfstoffen zugrunde liegt, ist einfach: Die Verabreichung eines harmlosen Derivats eines Infektionserregers soll das Immunsystem zur Bildung von Antikörpern anregen, die gegen diesen Erreger schützen.
Die Einführung einer fremden Substanz in den Körper kann jedoch nie völlig risikofrei sein, so dass die wichtigste Frage lautet, ob der Nutzen die Risiken überwiegen dürfte. Deshalb muss der Erreger gefährlich sein – eine Infektion mit ihm ist mit einer hohen Morbiditäts- und Mortalitätsrate verbunden, und die Impfung muss einen robusten immunologischen Schutz gegen schwere Erkrankungen bieten.
Diese Voraussetzungen waren bei den historischen Erfolgen bei der Entwicklung von Impfstoffen gegen Pocken, Tetanus, Diphtherie und Poliomyelitis erfüllt. In der Euphorie über diese wissenschaftlichen Meilensteine wurde jedoch eine entscheidende Tatsache übersehen. In allen vier Fällen wurden die Erreger in der Blutbahn an ihren Bestimmungsort transportiert, wo sie von den Antikörpern abgefangen werden konnten.
Es ist wichtig zu wissen, dass dies die Ausnahme und nicht die Regel ist. Die meisten viralen Erreger verursachen selbstlimitierende Infektionen der Atemwege oder des Magen-Darm-Trakts. Schwere Schäden an inneren Organen, die durch ihre Verbreitung über den Blutkreislauf verursacht werden, treten nur selten auf, und die Infektionen sind im Allgemeinen nicht mit hohen Sterberaten verbunden. Aufgrund ihrer Allgegenwärtigkeit ist in der Bevölkerung bereits eine hohe Grundimmunität gegen diese Viren vorhanden. Aus diesen einfachen Gründen besteht kein echter Bedarf an der Entwicklung von Impfstoffen gegen die meisten viralen Krankheitserreger.
Übrigens: Wenn Sie solche Informationen interessieren, dann fordern Sie unbedingt meinen kostenlosen Praxis-Newsletter dazu an:
Immunität gegen Atemwegsviren: systemische versus mukosale Immunität
Wir kommen nun zu einer wichtigen Tatsache, die den Schutz der Atemwege vor Infektionen betrifft: Er wird durch Zellen des Immunsystems vermittelt, die sich in und unter unseren Atemwegsschleimhäuten befinden; und diese Zellen funktionieren völlig unabhängig von den Immunzellen, die unsere inneren Organe schützen.
Ein Schlüsselaspekt dieser funktionellen Trennung zwischen mukosaler und systemischer Immunität betrifft die Art der Antikörper, die von den direkt unter den Schleimhäuten befindlichen Plasmazellen produziert werden. Diese Antikörper – das sekretorische Immunglobulin A (sIgA) – werden über die Schleimhäute an deren Oberfläche sezerniert. Sie sind also vor Ort, um auf Viren zu treffen, die über die Luft übertragen werden, und können sie daran hindern, sich an die Zellen in diesen Schleimhäuten zu binden und sie zu infizieren. Die gleiche Art des Schutzes gilt auch für den Verdauungstrakt.
Im Gegensatz dazu sind IgG und zirkulierendes IgA die wichtigsten Antikörper, die im Blutkreislauf zu finden sind. Sie können das Eindringen von Viren in die Zellen, die die Atemwege oder den Darm auskleiden, nicht verhindern, und sie können bestenfalls ihrer Verbreitung entgegenwirken, wenn sie in den Blutkreislauf gelangen. Entscheidend ist, dass Impfstoffe, die in den Muskel – also in das Innere des Körpers – injiziert werden, nur IgG und zirkulierendes IgA, nicht aber sekretorisches IgA induzieren. Die durch solche Impfstoffe induzierten Antikörper können und werden daher die Zellen des Respirationstraktes nicht wirksam vor einer Infektion durch luftübertragene Viren schützen [1,2]. Diese Erkenntnis ist weder umstritten noch neu. Bereits vor 30 Jahren kamen McGhee et al. [2] zu dem Schluss:
Es ist erstaunlich, dass trotz unseres heutigen Wissensstandes über das allgemeine Schleimhautimmunsystem fast alle aktuellen Impfstoffe dem Menschen auf parenteralem Weg [d. h. durch Injektion] verabreicht werden. Die systemische Immunisierung ist im Wesentlichen unwirksam für die Induktion von Schleimhautimmunreaktionen. Da die meisten infektiösen Mikroorganismen über die Schleimhäute aufgenommen werden, ist es logisch, die Induktion von schützenden Antikörpern und T-Zell-Reaktionen in den Schleimhäuten zu berücksichtigen.
Dass die intramuskuläre Injektion keine sekretorischen IgA induzieren kann, wurde in einer Studie zum Middle East Respiratory Syndrome (MERS) bestätigt [3]. Wie COVID-19 wird auch diese Krankheit durch ein Coronavirus verursacht, und der in der Studie verwendete experimentelle Impfstoff basierte auf einem Gen, wie alle wichtigen Impfstoffe, die derzeit gegen COVID-19 eingesetzt werden. Kürzlich hat eine andere Studie gezeigt, dass die mRNA-COVID-Impfstoffe ebenfalls keine substanzielle Produktion von sekretorischem IgA stimulieren [4]. Aus diesem einfachen Grund kann man nicht erwarten, dass eine Impfung die Infektion der Atemwege hemmt. In der Tat ist das völlige Versagen der Impfstoffe bei der Verhinderung einer SARS-CoV-2-Infektion heute solide dokumentiert [5,6].
Es ist allgemein bekannt, dass sekretorische IgA-Antikörper (sIgA) als Reaktion auf natürlich auftretende Infektionen der Atemwege gebildet werden. Die Schleimhäute gesunder Menschen sind folglich mit Antikörpern gegen häufige Atemwegsviren überzogen. Die Fähigkeit dieser Antikörper, Infektionen zu verhindern, ist jedoch begrenzt. Das Ergebnis einer Begegnung mit einem Virus ist nicht „schwarz oder weiß“ – es kommt auf die Zahlen an. Ein Schutzwall aus Antikörpern kann einen kleinen Angriff abwehren, wird aber bei einer höheren Viruslast überwunden. Daran ändert auch die Verwendung intranasaler Impfstoffe zur Stimulierung der sIgA-Produktion nichts, obwohl die intranasale Impfung eine stärkere mukosale Immunantwort auslöst als die intramuskuläre Injektion [3,7].
Die untergeordnete Rolle des sekretorischen IgA bei der Bekämpfung systemischer Virusinfektionen wird durch die Tatsache unterstrichen, dass Personen mit einem sehr häufigen genetischen Defekt – dem selektiven sIgA-Mangel -, die nicht in der Lage sind, sIgA zu produzieren, nicht unter einer dramatisch erhöhten Anfälligkeit für schwere Atemwegsinfektionen leiden. Diese Beobachtung lässt sich aus den folgenden beiden Prinzipien ableiten: Erstens beruht der immunologische Schutz gegen Atemwegsviren hauptsächlich auf T-Zellen, und zweitens sind bei Personen mit bereits bestehender Immunität die Antikörperspiegel im Blut (zirkulierendes IgG und IgA) im Allgemeinen ausreichend, um eine schwere Erkrankung durch Virusausbreitung im Körper zu verhindern.
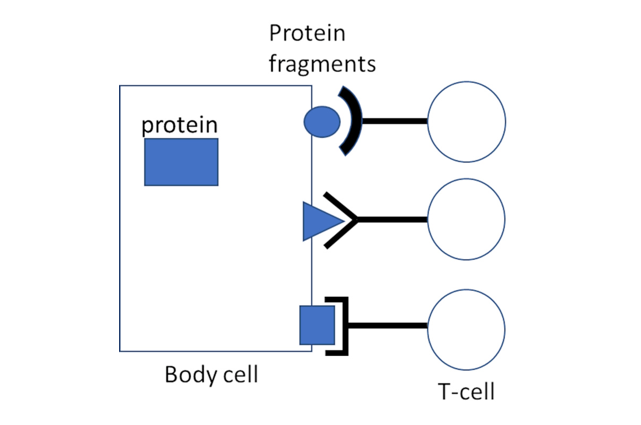
Abbildung 1: Schlüssel-Schloss-Interaktion zwischen Proteinfragmenten auf der Oberfläche einer Zelle und T-Zell-Rezeptoren zytotoxischer T-Zellen. Die Fragmente werden den T-Zellen durch ein spezifisches Trägermolekül, MHC 1 (nicht dargestellt), präsentiert. Die T-Zell-Rezeptoren auf den T-Lymphozyten unseres Körpers können insgesamt ein sehr breites Spektrum von Proteinfragmenten erkennen, aber alle Rezeptormoleküle auf einer bestimmten T-Zelle sind identisch und binden an dieselben Fragmente. T-Zellen, die an eines der von einem MHC-1-Molekül auf der Zelloberfläche präsentierten Proteinfragmente binden, werden dadurch aktiviert.
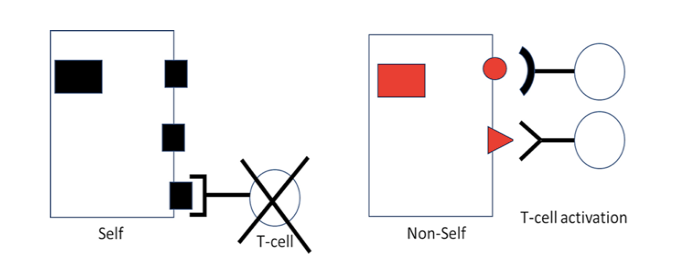
Wenn das Protein, dessen Fragmente diese CTL angelockt und aktiviert haben, von einem Virus kodiert wurde, dann wird das Ergebnis die Zerstörung der virusinfizierten Zelle sein, was für die Ausrottung einer Virusinfektion nützlich und notwendig ist. Es ist jedoch zu beachten, dass der Prozess der Proteinfragmentierung und -präsentation völlig allgemein ist – er ist nicht auf virale oder andere „Nicht-Selbst“-Proteine beschränkt, sondern gilt auch für körpereigene „Selbst“-Proteine. Daher ist es von entscheidender Bedeutung, die Aktivierung von CTL zu verhindern, die die Fragmente dieser aus „eigenen“ Proteinen gewonnenen Fragmente erkennen.
Übrigens: Wenn Sie solche Informationen interessieren, dann fordern Sie unbedingt meinen kostenlosen Praxis-Newsletter dazu an:
Wie wird dies bewerkstelligt?
Stellen Sie sich die Interaktion zwischen dem vorgestellten Proteinfragment und seinem „Rezeptor“ auf der T-Zelle als eine Interaktion zwischen Schloss und Schlüssel vor. Es gibt unzählige verschiedene Schlüssel (Fragmente), die in unzählige verschiedene Schlösser (T-Zell-Rezeptoren) passen. Es ist bekannt, dass die wahrhaft unglaubliche Vielfalt der Schlösser bereits während der fötalen Entwicklung entsteht.
Wie geschieht das? Werden die Schlösser als Reaktion auf die Fragmente (Schlüssel) geformt, wie sie während der Entwicklung erscheinen?
Da der Fötus in der Regel keinen Virusinfektionen ausgesetzt ist, wären die CTL mit Rezeptoren ausgestattet, die ausschließlich „eigene“ Proteinfragmente erkennen; aber diese selbstreaktiven CTL-Klone könnten kaum einem nützlichen Zweck dienen. Wenn hingegen die Vielfalt der Schleusen zufällig und ohne eine instruierende Vorlage (Schlüssel) entstehen sollte, dann müssten Milliarden von Lymphozyten, die „Nicht-Selbst“ erkennen – fremde Agenzien, einschließlich Virusproteine – neben denen entstehen, die „Selbst“ erkennen.
Erstaunlicherweise ist letzteres heute der Fall [8]. Auf wundersame Weise werden Lymphozyten, die „sich selbst“ erkennen, während des gesamten Lebens zum Schweigen gebracht oder in Schach gehalten, so dass sie gesunde Körperzellen nicht mutwillig angreifen können. Gelegentlich kommt es zu Missgeschicken, die zu einer Autoimmunerkrankung führen können. Wenn Zellen aus der Deckung kommen, die gegen Leberproteine reagieren, entsteht eine Autoimmunhepatitis. Fallen T-Zellen aus der Deckung, die auf die Inselzellen der Bauchspeicheldrüse reagieren, kommt es zu Autoimmun-Diabetes.
Andererseits sind die von Geburt an vorhandenen Immunzellen, die gegen praktisch alle Nicht-Selbst-Proteine reagieren, bereit, bei jeder Herausforderung in Aktion zu treten. Genau aus diesem Grund können herkömmliche Impfungen bereits im frühen Säuglingsalter erfolgreich durchgeführt werden. Und wenn ein Coronavirus auftaucht, erhebt sich das Anti-Corona-CTL-Team. Wenn eine Grippe auftritt, steigt das Anti-Grippe-Team auf, usw. Jedes Training stärkt das Team, so dass der Gegner schneller in Schach gehalten und Infektionen mit zunehmender Wirksamkeit beendet werden können.
Abbildung 2: Klonale Selektion von T-Lymphozyten. Die Vielfalt der T-Zell-Rezeptoren wird anfangs nach dem Zufallsprinzip erzeugt, was bedeutet, dass viele T-Zellen Rezeptoren tragen, die an Selbstantigene binden. Im Thymus werden solche T-Zellen durch Zellen, die diese Antigene exprimieren, „geködert“ und dann zerstört oder unterdrückt. T-Zellen, die keine Selbstantigene binden, bleiben bestehen und können zu einem späteren Zeitpunkt als Reaktion auf eine Virusinfektion aktiviert und zur Vermehrung angeregt werden.
Übrigens: Wenn Sie solche Informationen interessieren, dann fordern Sie unbedingt meinen kostenlosen Praxis-Newsletter dazu an:
Aber wird eine solche erworbene Immunität nicht durch immer neue „besorgniserregende“ Virusvarianten zunichte gemacht und unterlaufen?
Ganz im Gegenteil. Hier muss man beachten, dass ein Protein viele Fragmente erzeugt, die von vielen verschiedenen CTL-Klonen erkannt werden.
Die von einer Virusmutante kodierten Proteine können ein oder einige wenige unterschiedliche Fragmente erzeugen, aber die Mehrheit der anderen Fragmente wird gleich bleiben. Aus diesem Grund besteht eine CTL-basierte Kreuzreaktivität und ein Kreuzschutz zwischen allen Mitgliedern einer bestimmten Virusfamilie.
Speziell im Zusammenhang mit COVID-19 wurde festgestellt, dass bereits infizierte Personen in der Tat manchmal eine weitere Infektion mit einer neuen Variante durchmachen können, aber solche Reinfektionen sind fast nie ernsthafter Natur [9,10]. Das ist genau so, wie wir es hätten erwarten sollen; das Narrativ, dass das Auftreten von Virusmutationen durch die Entwicklung von maßgeschneiderten Impfstoffen bekämpft werden muss, war also von Anfang an grundlegend falsch.
Die Aktivierung von T-Lymphozyten – in diesem Fall jedoch von T-Helferzellen und nicht von CTL – ist auch an die Aktivierung von B-Lymphozyten gekoppelt, was zur Produktion von Antikörpern führt (Abbildung 3). Während CTL Fragmente von Proteinen erkennen, die auf der Zelloberfläche präsentiert werden, binden Antikörper an die intakten Proteine selbst.
Gebundene Antikörper lösen dann die Aktivierung eines anderen wichtigen Teils der Immunabwehr, des Komplementsystems, aus, was weitreichende Folgen hat. Durch die Aktivierung des Komplementsystems wird eine Vielzahl von Entzündungsprozessen ausgelöst. Außerdem greift das Komplementsystem selbst die Zelle an, auf deren Oberfläche die Aktivierung stattfindet, und zerstört diese.
Jeder genbasierte Impfstoff „encoding non-self“ ist äußerst gefährlich
Daraus folgt, dass die Produktion von „Nicht-Selbst“-Antigenen durch unsere eigenen Körperzellen unweigerlich entzündliche und zellzerstörende Prozesse auslöst. Bei Virusinfektionen ist dies sinnvoll, weil es zur Eliminierung der befallenen Zellen führt. Die meisten Viren greifen nur ein begrenztes Spektrum von Geweben an, und die meisten Gewebe können sich regenerieren, so dass Wunden anschließend heilen können.
Befürworter gentechnisch hergestellter Impfstoffe argumentieren häufig, dass diese Mittel nichts anderes tun, als das nachzuahmen, was bei tatsächlichen Virusinfektionen passiert. So wird behauptet, dass die Expression des fremden Proteins nur von kurzer Dauer ist und sich hauptsächlich auf die Stelle der intramuskulären Injektion beschränkt. Auch die Zellschädigung soll sich in Grenzen halten, so dass ernsthafte Nebenwirkungen nicht zu erwarten sind.
Nichts könnte irreführender und weiter von der Wahrheit entfernt sein.
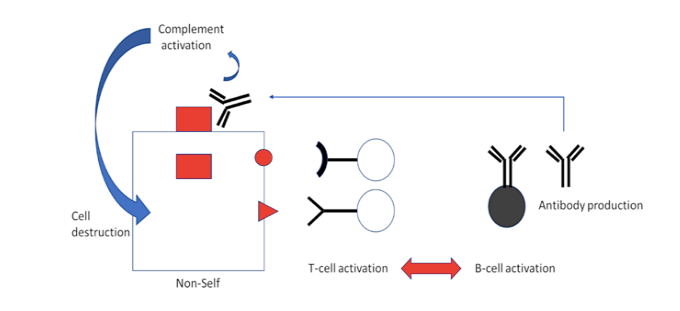
Abbildung 3: Zusammenarbeit von T-Zellen und Antikörpern bei der antiviralen Abwehr. T-Helferzellen werden durch die Fragmente eines viralen „Nicht-Selbst“-Antigens in ähnlicher Weise aktiviert wie die CTL. Ihre Aufgabe besteht jedoch nicht darin, selbst zum Angriff überzugehen; stattdessen aktivieren sie ihrerseits B-Zellen, die dann Antikörper gegen das intakte Nichtselbst-Protein produzieren. Wenn diese Antikörper ihr Ziel auf der Oberfläche einer infizierten Zelle finden, aktivieren sie das Komplement, eine Kaskade von Serumproteinen, die diese Zelle zerstören und auch die Entzündung im Allgemeinen fördern können.
Die Behauptung, dass die in LNP verpackte mRNA an der Injektionsstelle verbleibt, ist inzwischen weithin bekannt und stellt eine eklatante Unwahrheit dar. Diese „Impfstoffe“ verbreiten sich rasch von der Injektionsstelle zu den Lymphknoten und in den Blutkreislauf [11], und eine langlebige Expression in Organen und Geweben in einiger Entfernung von der Injektionsstelle wurde wiederholt und mit einer Reihe von Analyseverfahren dokumentiert [12-15]. Und da die Impfstoffpartikel in alle kernhaltigen Zellen eindringen können, wird ihre Aufnahme in Zellen der Lymphknoten, in Endothelzellen, die die Wände der Blutgefäße auskleiden, und in Zellen jedes Gewebes, das sie erreichen, zwangsläufig rasch erfolgen.
Diese Tatsache unterscheidet die „mRNA-Impfung“ sofort von natürlich vorkommenden Infektionen. Nur sehr wenige Infektionserreger greifen systemisch Lymphozyten oder Endothelzellen an. Zu letzteren gehören gefährliche Viren, die hämorrhagisches Fieber verursachen, und Bakterien, die ebenfalls lebensbedrohliche Infektionen hervorrufen, z. B. Typhus und Rocky Mountain Fleckfieber.
In krassem Gegensatz dazu löst jeder einzelne mRNA-„Impfstoff“ selbstzerstörerische Prozesse in den lymphatischen Organen und in den Blutgefäßen des Körpers aus. Die immensen Gefahren von Selbstangriffsereignissen innerhalb des immunologischen Kontrollnetzes sind bereits beschrieben worden [16]. Dazu gehören die Reaktivierung ruhender Infektionen (z. B. Herpes simples, Gürtelrose, EBV, CMV, Tuberkulose, Parasiten), eine verringerte Fähigkeit zur Kontrolle neuer Infektionen und die Aktivierung oder Reaktivierung von Neoplasmen [17].
Gleichzeitig findet ein konzertierter Immunangriff gegen die Gefäßwände statt, wann immer und wo immer die Endothelzellen transfiziert werden (Abbildung 4). Im Falle von SARS-CoV-2 ist bekannt, dass Spike-Protein-spezifische zytotoxische T-Zellen im Blut gesunder Personen weit verbreitet sind. Dies kann auf eine frühere Infektion mit diesem Virus zurückzuführen sein, aber auch auf eine immunologische Kreuzreaktivität mit anderen, verwandten Coronaviren [18,19]. Mit dem Auftreten spezifischer Antikörper wird der Angriff auf Zellen, die die fremden Proteine tragen, vervielfacht und durch die Wirkung von Komplement und phagozytischen Zellen intensiviert. Blutgerinnsel, die sich im Gefolge der Endothelverletzung bilden, führen zu Durchblutungsstörungen. Der ischämische Zelltod hat irreversible Folgen für das zentrale Nervensystem und das Herz. Schäden an den Gefäßwänden führen vorhersehbar zum Austritt von Impfstoffen in die Zellen der betreffenden Organe und zu deren Aufnahme durch sie, was den Keim für unzählige autodestruktive Ereignisse legt.
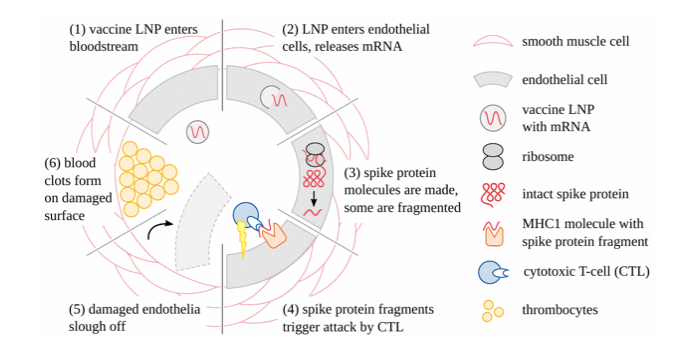
Abbildung 4: Wie COVID-19 mRNA-Impfstoffe die Blutgefäße schädigen und die Blutgerinnung verursachen. Nachdem die Lipid-Nanopartikel des Impfstoffs in den Blutkreislauf gelangt sind, werden sie von den Endothelzellen aufgenommen, und die mRNA wird freigesetzt. Das Spike-Protein wird dann exprimiert; einige Moleküle werden fragmentiert und von einem speziellen Trägerprotein (MHC1) auf der Zelloberfläche präsentiert. Dies führt dazu, dass die Endothelzellen von zytotoxischen T-Zellen angegriffen werden. Die zerstörten Endothelzellen schälen sich ab, wodurch die Impfstoffpartikel in das angrenzende Gewebe gelangen können. Dadurch werden auch die tieferen Schichten der Gefäßwand dem Blut ausgesetzt, was die Thrombozytenaggregation und die Blutgerinnung auslöst.
Die sich häufenden Daten bestätigen diese Vorahnungen, und ein neuer Befund könnte sich als charakteristisch und diagnostisch für impfstoffvermittelte Pathologien erweisen: Die impfstoffinduzierte Expression von Spike-Protein in Endothelzellen und die daraus resultierende Vaskulitis gehen Hand in Hand. Die erste Illustration dieses Prinzips wurde in einem Fallbericht über einen 76-jährigen Mann vorgestellt, der drei Wochen nach seiner dritten COVID-19-Impfung verstarb [20]. Die histopathologischen Untersuchungen des Gehirns ergaben eine multifokale Vaskulitis und eine nekrotisierende Enzephalitis. Im Herzen wurden eine Vaskulitis der kleinen Gefäße und eine lymphozytäre Myokarditis festgestellt. Spike-Protein wurde innerhalb der Entzündungsherde sowohl im Gehirn als auch im Herzen nachgewiesen, insbesondere in den Endothelzellen der kleinen Blutgefäße (Abbildung 5). Entsprechende Kontrollexperimente bestätigten, dass die beobachtete Spike-Protein-Expression tatsächlich durch die Impfstoffinjektionen verursacht wurde, die der Patient erhalten hatte, und nicht durch eine nicht diagnostizierte Infektion mit dem Virus selbst.
Multiorganische Vaskulitis, bei der vor allem kleine Gefäße betroffen sind, ist ein häufiges Thema bei den unerwünschten Ereignissen nach der COVID-19-Impfung. Der Befall von Kapillaren mit Thrombusbildung in Gehirn und Herz wird immer wieder beobachtet (Mörz [20]; Mörz und Burkhardt, unveröffentlicht). Das beobachtete Krankheitsbild in kleinen und kleinsten Gefäßen ist neu und wird von den Autoren als charakteristisch für die Wirkung von genbasierten Impfstoffen angesehen.
Fulminante Reaktionen sind bei Patienten zu erwarten, die nach Erholung von einer echten SARS-CoV-2-Infektion geimpft werden. Solche Patienten haben hohe Konzentrationen an zirkulierenden IgG-Antikörpern gegen das Spike-Protein [21], und der Komplementangriff auf impfstofftransfizierte Zellen kann dann sofort und massiv sein. Es wurde ein Fallbericht über einen durch Myokarditis ausgelösten plötzlichen Tod nach der ersten Impfung mit direktem Nachweis einer Komplementaktivierung im Herzen veröffentlicht [22].
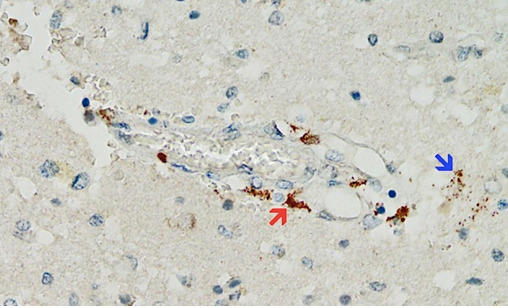
Abbildung 5: Expression von SARS-CoV-2-Spike-Protein im Gehirn eines dreifach geimpften Patienten (Immunhistochemie). Das braune Pigment zeigt das Spike-Protein in einem kleinen Blutgefäß (roter Pfeil) sowie in Gliazellen des umgebenden Hirngewebes (blauer Pfeil). Das Foto stammt von Mörz [20].
Es stimmt, dass das Spike-Protein selbst die Blutgerinnung und Entzündung ohne Hilfe des Immunsystems fördern kann [23]. Dennoch deuten die bereits vorliegenden Beweise darauf hin, dass die schwerwiegenden, weit verbreiteten und anhaltenden Schäden an Geweben und Blutgefäßen hauptsächlich durch den Angriff des Immunsystems auf die Spike-Protein produzierenden Zellen verursacht werden.
Dieser Angriff erfolgt einfach deshalb, weil das Spike-Protein ein Nicht-Selbst-Antigen ist; und da jeder andere mRNA-Impfstoff für sein eigenes Nicht-Selbst-Antigen kodiert, das von der jeweiligen Mikrobe stammt, auf die er abzielt, müssen wir davon ausgehen, dass er durch denselben Mechanismus und in ähnlichem Umfang Schaden anrichten wird. Diese alptraumhaften Szenarien werden mit jeder Auffrischungsimpfung nur noch schlimmer werden. Die katastrophalen Ereignisse werden aufgrund ihrer Natur weder vermeidbar noch unterdrückbar sein.
Die Katastrophe, die sich vor unseren Augen abspielt, könnte und wurde [24] von den ersten Prinzipien der Immunologie vorhergesagt. Die Fähigkeit, zwischen Selbst und Nicht-Selbst zu unterscheiden, ist grundlegend für das Leben. Sie ist bereits bei der Geburt vorhanden und endet erst mit dem Tod. Sie kann nicht manipuliert oder kontrolliert werden. Jeder Versuch, dies mit mRNA oder anderen genbasierten Impfstoffen zu tun, ist zum Scheitern verurteilt.
Übrigens: Wenn Sie solche Informationen interessieren, dann fordern Sie unbedingt meinen kostenlosen Praxis-Newsletter dazu an:
Quellen:
Der Beitrag ist eine Übersetzung des Originals unter:
https://www.globalresearch.ca/gene-based-vaccination-quo-vadis/5797968
Die angegebenen Quellen im Beitrag:
- Kurono, Y. (2021) The mucosal immune system of the upper respiratory tract and recent progress in mucosal vaccines. Auris nasus larynx (preprint)
- McGhee, J.R. et al. (1992) The mucosal immune system: from fundamental concepts to vaccine development. Vaccine 10:75-88
- Kim, M.H. et al. (2019) Superior immune responses induced by intranasal immunization with recombinant adenovirus-based vaccine expressing full-length Spike protein of Middle East respiratory syndrome coronavirus. PLoS One 14:e0220196
- Meyer-Arndt, L. et al. (2022) Cutting Edge: Serum but Not Mucosal Antibody Responses Are Associated with Pre-Existing SARS-CoV-2 Spike Cross-Reactive CD4+T Cells following BNT162b2 Vaccination in the Elderly. Immunol. 208:1001-1005
- Chau, N.V.V. et al. (2021) Transmission of SARS-CoV-2 Delta Variant Among Vaccinated Healthcare Workers, Vietnam.
- Singanayagam, A. et al. (2021) Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: a prospective, longitudinal, cohort study. Lancet Infect. Dis. (preprint)
- Du, L. et al. (2008) Intranasal vaccination of recombinant adeno-associated virus encoding receptor-binding domain of severe acute respiratory syndrome coronavirus (SARS-CoV) spike protein induces strong mucosal immune responses and provides long-term protection against SARS-CoV infection. Immunol. 180:948-56
- Rechavi, E. and Somech, R. (2017) Survival of the fetus: fetal B and T cell receptor repertoire development. Immunopathol. 39:577-583
- Dhar, M.S. et al. (2021) Genomic characterization and epidemiology of an emerging SARS-CoV-2 variant in Delhi, India. Science 374:995-999
- Altarawneh, H. et al. (2022) Protection afforded by prior infection against SARS-CoV-2 reinfection with the Omicron variant. medRxiv (preprint)
- Anonymous, (2020) SARS-CoV-2 mRNA Vaccine (BNT162, PF-07302048) 2.6.4 Summary statement of the pharmacokinetic study [English translation].
- Bansal, S. et al. (2021) Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer-BioNTech) Vaccination prior to Development of Antibodies: A Novel Mechanism for Immune Activation by mRNA Vaccines. Immunol. 207:2405-2410
- Magen, E. et al. (2022) Clinical and Molecular Characterization of a Rare Case of BNT162b2 mRNA COVID-19 Vaccine-Associated Myositis. Vaccines 10 (preprint)
- Röltgen, K. et al. (2022) Immune imprinting, breadth of variant recognition and germinal center response in human SARS-CoV-2 infection and vaccination. Cell (preprint)
- Yamamoto, M. et al. (2022) Persistent varicella zoster virus infection following mRNA COVID‐19 vaccination was associated with the presence of encoded spike protein in the lesion. Cutan. Immunol. Allergy (preprint)
- Anonymous, (2021) Shots and Shingles: What Do They Tell Us?.
- Krüger, U. (2022) COVID vaccination and turbo cancer: pathological evidence.
- Grifoni, A. et al. (2020) Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 181:1489-1501.e15
- Nelde, A. et al. (2020) SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nature immunology (preprint)
- Mörz, M. (2022) A Case Report: Multifocal Necrotizing Encephalitis and Myocarditis after BNT162b2 mRNA Vaccination against Covid-19. Vaccines 10:2022060308
- Killingley, B. et al. (2022) Safety, tolerability and viral kinetics during SARS-CoV-2 human challenge in young adults. Med. (preprint)
- Choi, S. et al. (2021) Myocarditis-induced Sudden Death after BNT162b2 mRNA COVID-19 Vaccination in Korea: Case Report Focusing on Histopathological Findings. Korean Med. Sci. 36:e286
- Letarov, A.V. et al. (2021) Free SARS-CoV-2 Spike Protein S1 Particles May Play a Role in the Pathogenesis of COVID-19 Infection. Biochemistry Mosc 86:257-261
- Bhakdi, S. et al. (2021) Urgent Open Letter from Doctors and Scientists to the European Medicines Agency regarding COVID-19 Vaccine Safety Concerns.

